
.png)
.png)

了解步态发育对于更好地识别和治疗儿童早期运动迟缓很重要。恒河猴步态有着宽阔的支撑基础和较短的步伐来弥补不稳定性,随着幼年过渡到更成熟、变异性更小的步态模式,他们在步态周期中的平衡和控制能力会增强。步态已被用作一种衡量标准,可以根据这些步态参数将神经发育障碍与典型发育区分开,如自闭症谱系障碍儿童的运动缺陷、注意力缺陷/多动障碍、脑瘫、发育协调障碍,以及一般的智力和发育残疾。
恒河猴(猕猴)是研究运动发育的理想模型,与其他动物模型相比,它们更接近人类胎儿大脑。它们的发育速度也是人类的3-4倍,研究恒河猴的步态是治疗脊髓损伤、帕金森氏症和亨廷顿舞蹈症的有效方法。通过将恒河猴模型扩展到神经发育研究中,可以精确定位步态发育的细微差异,并将其作为人类运动障碍的重要标志。
本研究将诺达思的动物步态分析系统(CatWalk)应用到恒河猴创建步态的客观测量。动物步态分析系统最初设计用于测量啮齿动物的步态,并已成功应用于成年普通狨猴和灰鼠狐猴非人灵长类动物模型。本研究的目的是检验动物步态分析系统收集幼年恒河猴步态数据的可行性,并量化出生后第一个月的初步步态发展。
2.1数据收集
选择12只印度裔恒河猴(Macaca mulatta),使用施耐德灵长类新生儿评估 (SNAP) 和动物步态分析系统进行神经发育测试。首先是 SNAP 测试,大约需要 30 分钟来测试恒河猴的运动、定向、感觉反应和状态控制。恒河猴在SNAP运动测试中的表现用于进一步了解恒河猴使用动物步态分析系统完成步态评估的能力。
动物步态分析系统为了更好针对幼年恒河猴研究进行了调整。研究小组成员直接将恒河猴抱起并放入动物步态分析系统的入口处,恒河猴在130厘米长的走道中走动(图 1)。 当动物穿过玻璃走道并对玻璃走道施加压力时,绿色 LED 会折射到红色背景上,高速数码相机会捕捉到每只动物的脚步。 在走道的尽头,恒河猴进入目标箱。
每只恒河猴的每部分肢体必须在走道上至少有两次连续的脚步,不得停止或跳跃。对恒河猴进行测试,直到收集到至少三个可用的运行数据,或者恒河猴无法或不愿意完成任务。如果恒河猴没有完成可用的前进,测试人员会尝试通过让恒河猴部分穿过动物步态分析系统系统或将他们的毯子放在系统中作为强化物来训练恒河猴。 数据在出生后第14天、第21天和第28±1天收集。恒河猴的出生被归类为生命的第一天。
2.2 数据准备
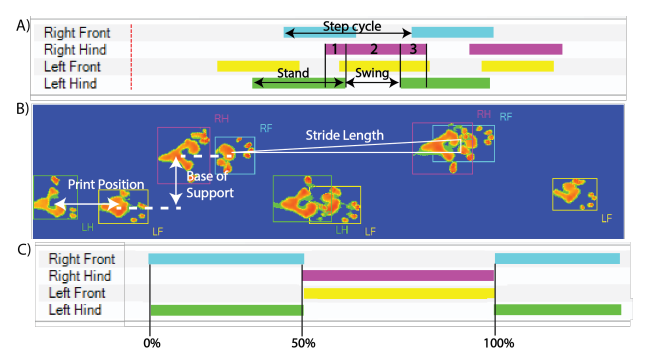
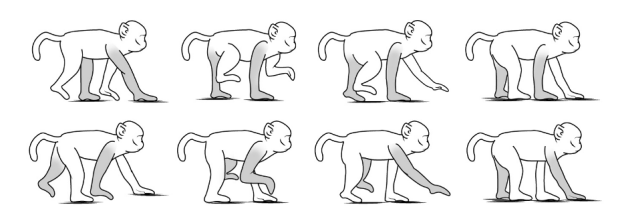
所有肢体都标记为右前 (RF)、左前 (LF)、右后 (RH) 和左后 (LH)。动物步态分析系统计算了所有变量在每个时间点的统计数据,即三次前进的平均值。我们选择动物步态分析系统中使用步态和爪子变量,这些变量被用于测量平衡和协调的发展。选择的变量分为速度、动态爪测量、静态爪测量和爪间协调。表2中列出了选定的变量,包括定义、肢体输出和方程(图2)。当玻璃走道的所有四个肢体都可见时,计算开始,当至少一个肢体不再可见时结束。
2.3行走方式分类
为了确保客观的步态数据反映行走模式,对所有恒河猴的行走方式进行了归类:对角线或非对角线模式。对角线行走模式被定义为在摆动和站立阶段,对侧前肢和后肢及时靠近(图3)。非对角线模式被定义为任何其他肢体组合,包括当同侧前肢和后肢在时间上摆动并靠得很近时,以及如果观察到对角线和非对角线模式的混合组合。
3.1 动物步态分析系统的可行性和可靠性
评估12只动物完成至少一次穿越步道的能力(每个肢体在走道上至少连续两次脚步而不停止或跳跃)。恒河猴执行任务的能力在第14天时最低 (66.7%),在第21天时有所增加 (75%),到第28天时,大多数恒河猴 (83.3%) 至少有一次可用的前进。在这些不同时间点能够完成至少一次前进的恒河猴比例没有显著差异(表 2)。无法在第21天和第28天完成任务的恒河猴会抵制进入动物步态分析系统设备或因环境而分心,而第14天的恒河猴似乎在运动技能方面存在困难。我们探索了一个时间点内每个恒河猴运行之间连续变量的重测可靠性(表3)。
3.2 步态数据
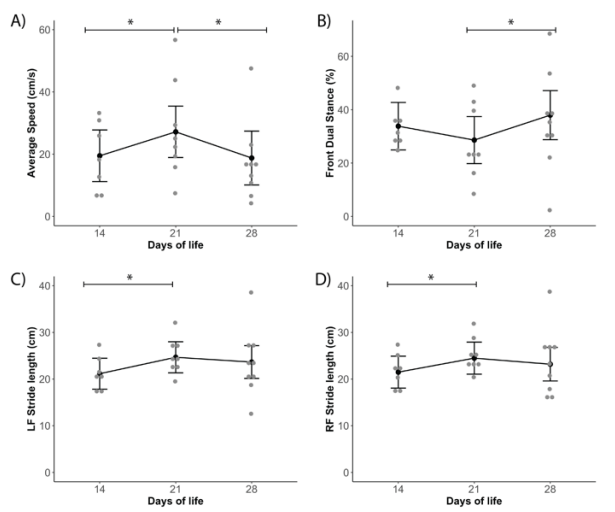
本研究的第二个目标是检查出生后 14 天、21 天和 28 天的步态参数差异。 在调整了恒河猴在与母亲一起安置之前的天数和出生体重后,我们计算了每天的平均值和 95% CI(表 3)。速度从第14天到第21天显著增加(p=0.049),然后从第21天到第28天(p=0.032)下降,第14天和第28天之间没有差异(图 4A)。 从第21天到第28天,恒河猴将双前爪保持在走道上(即双姿势)的时间百分比显著增加(p = 0.035),而从第14天到第21天没有明显变化(图 4B)。 从第14天到第21天,前步长显著增加(LF p=0.019;RF p=0.049),然后在第28天保持稳定(图 4C、D)。 在出生第14天至第28天期间,恒河猴在斜肢、三肢和四肢上花费的时间大致相同,在四肢站立方面没有显著差异。 每个时间点都存在广泛的变异性,如大范围的置信区间所示(表3)
3.3 行走模式
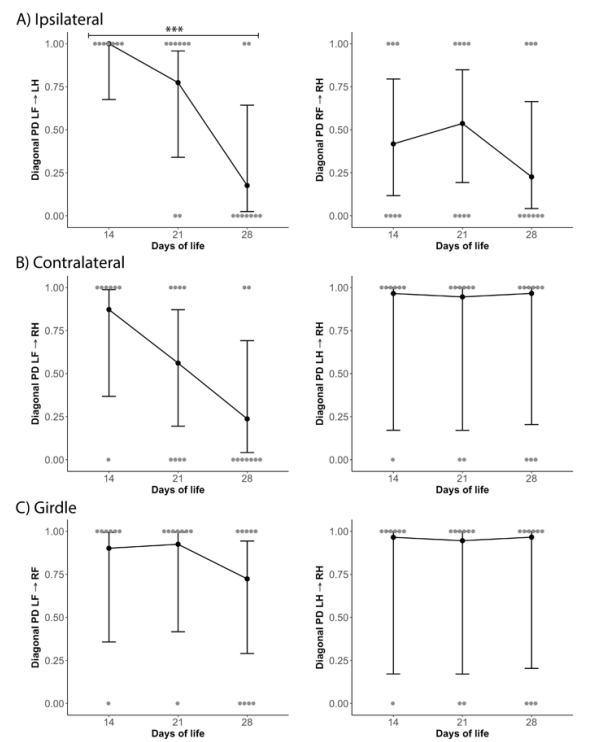
对角行走模式,即对侧前肢和后肢同步移动,被认为是恒河猴发育成熟的行走模式。 我们比较了相位色散参数(phase dispersion parameters),该参数代表锚点和目标肢体放置在走道上之间的时间。在成熟的对角行走模式中,我们预计相位色散参数对侧肢体对的 cstat 平均值为 0 或 100%,同侧和腰带肢体对为50%。 根据预期范围内的cstat平均情况,使用或不使用对角线模式,将相位色散转换为锚点和目标肢体的分类变量(图 5)。 我们发现 LF→ LH 存在一个显著差异,在生命第14天,100% 的肢体对计时落在对角相位分散内,而在生命第28天,只有 17.6% (p=0.0002)。 在生命的第 14 天,除了 RF→RH 之外的所有肢体对的时间都落在对角行走模式内。 这些发现表明,幼年恒河猴在第14天左右开始采用对角线行走模式。
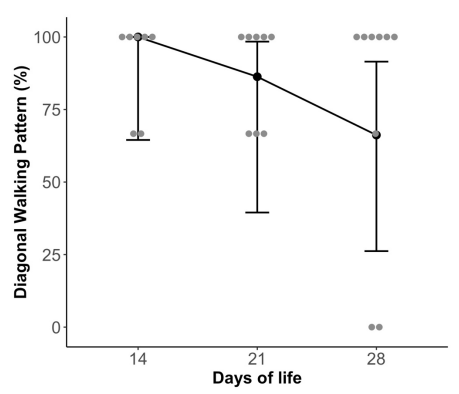
我们发现第14天、第21天和第28天之间采用对角行走模式的动物百分比没有显著差异(图 6)。50%的恒河猴在动物步态分析系统行走时,对角行走模式的时间安排略有不同。 前肢通常比后肢稍早落地,但在站立和摆动阶段仍然使用对侧肢体一起移动的对角模式。
本研究的目的是将动物步态分析系统在恒河猴模型中应用扩展,并定义出生后 14 天、21天和28天的初步规范步态发育。这项研究表明,使用动物步态分析系统收集幼年恒河猴出生第一个月的步态数据是可行的。该技术提供了典型步态的定量分类,显示出生后第一个月整体运动发育协调和平衡变量随时间的有限变化。
恒河猴在出生14天时就已经开始采用对角线行走模式。这与之前的文献形成鲜明对比。 有两只恒河猴在出生第 28 天时不符合对角行走模式的定义,它们被归类为奔跑或跳跃,这是更成熟的模式。 这些恒河猴可能会走得更快或探索更成熟的前进/奔跑模式,从而导致对角线模式未被利用。 在所有时间点,恒河猴的时间都有变化,大约一半的恒河猴使用对角线模式,前肢比后肢早几秒钟落地,时间略有延迟。这一发现可能表明,恒河猴早期已经采用对角行走模式,但可能具有可变的协调性,这可以通过移动对侧肢体的时间变化来证明。 这种差异的识别可能是使用动物步态分析系统技术的一个新颖结果,与之前的观察评估相比,该技术允许对恒河猴行走模式进行逐帧分析。
从第14天到第28天,我们没有发现步态参数出现显著的发育变化,这与我们最初的假设相反。这项研究发现,在第14天到第28天的发育过程中,相分散 LF→LH 只有一个显著变化。我们发现,在每个时间点,动物步态分析系统各个变量在动物内部和动物之间存在广泛的变异性。这可能进一步反映了幼年恒河猴的典型步态发育范围,这些数据与人类发育一致。
Kabakov, Sabrina A., et al. “Quantification of early gait development: Expanding the application of Catwalk technology to an infant rhesus macaque model.” Journal of Neuroscience Methods 388 (2023): 109811.





